Artemia franciscana
Artemia franciscana có thể được dùng để sản xuất axit béo thiết yếu?
Artemia, đặc biệt là giai đoạn đầu vòng đời của chúng (nauplii), được cho là con mồi sống được sử dụng phổ biến nhất trong nuôi ấu trùng hải sản do tiết kiệm chi phí, dễ xử lý và có nhiều giá trị.
Tuy nhiên, một hạn chế lớn liên quan đến việc sử dụng ấu trùng Artemia làm thức ăn là giá trị dinh dưỡng thiếu hụt đối với ấu trùng cá biển có liên quan chủ yếu đến mức độ dưới mức tối ưu của các axit béo không bão hòa đa chuỗi dài (LC-PUFA).
Thật vậy, LC-PUFA, bao gồm axit eicosapentaenoic (EPA, 20:5n-3), axit arachidonic (ARA, 20:4n-6) và axit docosahexaenoic (DHA, 22:6n-3), là những chất dinh dưỡng thiết yếu hỗ trợ sự tăng trưởng tối ưu và tỷ lệ sống của ấu trùng.
Trong khi một số loài cá có khả năng sinh tổng hợp LC-PUFA (Castro và cộng sự, 2016), các loài cá ăn thịt biển bao gồm cá tráp, cá vược, cá ngừ, hổ phách, v.v., phần lớn được coi là bị hạn chế trong khả năng sản xuất LC-PUFA nội sinh. (Xie và cộng sự, 2021; Monroig và cộng sự, 2022). Do đó, chế độ ăn cho cá biển phải chứa đủ hàm lượng LC-PUFA để đáp ứng yêu cầu về các chất dinh dưỡng thiết yếu này (NRC, 2011), với nhu cầu như vậy đặc biệt cao trong giai đoạn đầu phát triển khi các mô thần kinh tích tụ các hợp chất này đang hình thành nhanh chóng (Uauy et al., 2001; Janssen và Kiliaan, 2014).
Thành phần axit béo (FA) của ấu trùng Artemia mới nở thường được đặc trưng bởi có một số mức EPA, nhưng mức độ dấu vết hoặc không thể phát hiện được của DHA (Navarro et al., 1993). Tuy nhiên, hàm lượng EPA phần lớn khác nhau giữa các bào xác và ấu trùng có nguồn gốc địa lý khác nhau và điều này dẫn đến việc phân loại Artemia thành hai nhóm chính, đó là “Artemia loại biển” với hàm lượng EPA tương đối cao và axit α-linolenic (ALA) thấp, và “Artemia loại nước ngọt” có hàm lượng EPA tương đối thấp và ALA cao (Watanabe và cộng sự, 1978; Watanabe và cộng sự, 1980; Navarro và cộng sự, 1993). Mặc dù cách phân loại này đã được sử dụng rộng rãi, nhưng nguyên nhân cụ thể tạo ra sự khác biệt trong đặc điểm FA của Artemia vẫn chưa được biết rõ.
 Artemia franciscana được cho là loài Artemia được sử dụng phổ biến nhất trong nuôi trồng thủy sản trên toàn thế giới
Artemia franciscana được cho là loài Artemia được sử dụng phổ biến nhất trong nuôi trồng thủy sản trên toàn thế giới
Nghiên cứu mới đây của Marc Ramos-Llorens và cs (2023) được thực hiện nhằm mục đích tìm kiếm rộng rãi các gen tiềm năng liên quan đến quá trình sinh tổng hợp LC-PUFA ở Artemia franciscana, được cho là loài Artemia được sử dụng phổ biến nhất trong nuôi trồng thủy sản trên toàn thế giới. Bằng cách sử dụng các tài nguyên tin sinh học có sẵn cho A. franciscana (Jo và cộng sự, 2021), các tác giả đã thực hiện tìm kiếm có hệ thống các gen mã hóa Elovl, cũng như các desaturase đầu tiên, methyl-end và front-end, từ A. franciscana, và thực hiện đặc tính phân tử và chức năng của các enzyme được mã hóa.
Kết quả giải trình tự và phát sinh loài của elongase A. franciscana
Tổng cộng có tám trình tự giống Elovl (Elo1-8) đã được lấy từ cơ sở dữ liệu gen của A. franciscana và gửi vào NCBI GenBank với các số gia nhập MZ262329 (Elo1), MZ262324 (Elo2), MZ262325 (Elo3), MZ262326 (Elo4) , MZ262327 (Elo5), MZ262328 (Elo6), OM677865 (Elo7) và ON416870 (Elo8). ORF của bản mô tả này được gọi là Elo1 từ A. franciscana bao gồm 993 cặp bazơ (bp), mã hóa protein giả định có kích thước 330 aa. Hơn nữa, các chuỗi ORF của A. franciscana Elo2, Elo5, Elo6 và Elo7 lần lượt bao gồm 990, 897, 909 và 807 bp, mã hóa các protein giả định là 329, 298, 302 và 268 aa. Cả Elo3 và Elo4 đều có ORF là 1035 bp, mã hóa protein giả định là 344 aa. Cuối cùng, Elo8 bao gồm 864 bp mã hóa 287 aa.
Phân tích các trình tự aa được suy ra cho thấy rằng tất cả tám trình tự giống Elovl từ A. franciscana đều chứa hộp H đặc biệt (HXXHH) (Hình 1). Hơn nữa, các gốc aa nằm ở phía đầu N của hộp H, được sử dụng để phân biệt các elongase PUFA và không PUFA (Hashimoto và cộng sự, 2008), cho thấy một số khác biệt giữa sáu elongase A. franciscana. A. franciscana Elo1, Elo2, Elo5, Elo6 và Elo7 chứa glutamine (Q) ở vị trí −5 trước hộp H, trong khi Elo3 và Elo4 chứa histidine (H) ở vị trí −5 (Hình 1). Trong khi cả hai dư lượng thay thế (Q và H) đều đáp ứng các tiêu chí do Hashimoto et al. (2008) đối với các elongase PUFA, A. franciscana Elo8 chứa proline (P) ở vị trí −5 trước hộp H (Hình 1) và do đó bị loại khỏi xét nghiệm mô tả đặc tính chức năng tiếp theo.

Hình 1. Việc sắp xếp các chuỗi axit amin được suy ra của các cDNA giống A. franciscana elovl được nghiên cứu trong tài liệu này (Elo1, Elo2, Elo3, Elo4, Elo5, Elo6, Elo7 và Elo8), được thực hiện bằng Geneious Prime Software (v: 2022.1. 1) và các trình tự được căn chỉnh theo Clustal Omega (cài đặt mặc định) như được triển khai trong phần mềm (Sievers và cộng sự, 2011; Kearse và cộng sự, 2012). Hộp histidine mô-đun bảo tồn Elongase (HXXHH) (Hashimoto và cộng sự, 2008) được gạch chân và vị trí hộp −5 histidine được đánh dấu bằng một mũi tên.
Các trình tự aa của A. franciscana Elovl được so sánh với các elongase từ nhiều loại động vật bằng cách xây dựng một cây phát sinh gen (Hình 2). Kết quả cho thấy protein A. franciscana Elo1 được nhóm lại với các chỉnh hình elovl4 từ động vật có xương sống và động vật không xương sống bao gồm cả các loài giáp xác như H. azteca và D. magna (Hình 2). Những kết quả này xác nhận rằng A. franciscana Elo1 mới được nhân bản là một bản chỉnh hình của Elovl4. Một cụm được hỗ trợ tốt khác chứa A. franciscana Elo2, Elo3, Elo4, Elo5 và Elo6, cùng với các elongase từ động vật giáp xác như S. olivacea, E. marinus, T. californicus, v.v., cũng được xác định trên cây (Hình 2). 2). Nhánh này chứa các chuỗi thuộc về Elovl1 và Elovl7 từ nhiều loài động vật có xương sống (Hình 2). A. franciscana Elo7 được nhóm lại cùng với các chuỗi Elovl8 được đặc trưng từ các teleost (ví dụ: Danio rerio) (Hình 2). Cuối cùng, A. franciscana Elo8 tập hợp lại với các chuỗi giống Elovl6. Không có elongase nào trong số tám A. franciscana elongase được nhóm lại với các trình tự tạo thành một nhánh chứa PUFA elongase Elovl2 và Elovl5 từ động vật có xương sống và động vật thân mềm Elovl2/5 (Hình 2).

Hình 2. Cây phát sinh gen so sánh A. franciscana Elo1, Elo2, Elo3, Elo4, Elo5, Elo6, Elo7 và Elo8 với protein elongase từ các sinh vật khác. Cây được xây dựng bằng phương pháp khả năng tối đa (Jones và cộng sự, 1992) bằng phần mềm MEGA-X (Kumar và cộng sự, 2018). Các con số biểu thị tần số (%) mà cấu trúc liên kết cây trình bày được sao chép sau 1000 lần lặp (Felsenstein, 1985).
Trình tự và phát sinh loài của A. franciscana desaturase
ORF của ba loại A. franciscana desaturase được gọi là Des1, Des2 và Des3 lần lượt chứa 1005, 1029 và 1053 bp, mã hóa các protein giả định là 334, 342 và 350 aa (Hình 3). Các chuỗi desaturase mới được nhân bản từ A. franciscana đã được gửi vào NCBI GenBank với các số gia nhập sau: MZ254648 (Des1), MZ254649 (Des2) và MZ254650 (Des3). Ba trình tự chứa ba đặc tính hộp H riêng biệt của desaturase (Hashimoto và cộng sự, 2008). Cụ thể, hộp H của A. franciscana desaturase là HXXXXH (H-box1), HXXHH (H-box2) và HXXHH (H-box3) (Hình 3). Thành phần dư lượng trong H-box1 (HRLWXH), H-box2 (HRVHH) và H-box3 (HNXHH) phù hợp với chất khử bão hòa “Đầu tiên” (Hashimoto et al., 2008). Một cách nhất quán, phân tích phát sinh gen của chúng tôi cho thấy rằng các protein A. franciscana Des1-3 được nhóm lại với nhau với các chất khử bão hòa đầu tiên hầu hết được biểu thị bằng các chỉnh hình stearoyl-CoA desaturase từ nhiều loại động vật và khác xa với các phân họ desaturase khác bao gồm cả các chất khử bão hòa ở đầu methyl và đầu cuối (Hình 4).

Hình 3. Việc căn chỉnh các chuỗi protein A. franciscana Des1, Des2 và Des3 được thực hiện bằng Geneious Prime Software (v: 2022.1.1) và các chuỗi được căn chỉnh với Clustal Omega (cài đặt mặc định) như được triển khai trong phần mềm (Sievers et al., 2011 ; Kearse và cộng sự, 2012). Các hộp histidine được bảo tồn cho các chất khử bão hòa đầu tiên ([HXXXXH], [HXXHH] và [HXXHH]) (Hashimoto et al., 2008) được gạch. chân.

Hình 4. Cây phát sinh loài so sánh A. franciscana Des1, Des2 và Des3 với trình tự desaturase từ nhiều loài động vật. Trình tự từ các loại enzyme khác nhau, cụ thể là desaturase đầu tiên, desaturase đầu methyl và desaturase đầu cuối, đã được phân tích. Cây được xây dựng bằng phương pháp khả năng tối đa (Jones và cộng sự, 1992) bằng phần mềm MEGA-X (Kumar và cộng sự, 2018). Các con số biểu thị tần số (%) mà cấu trúc liên kết cây trình bày được sao chép sau 1000 lần lặp (Felsenstein, 1985).
Vai trò của A. franciscana elongase trong quá trình sinh tổng hợp LC-PUFA
Khả năng kéo dài PUFA của các enzyme giống A. franciscana Elovl đã được kiểm tra bằng cách ủ nấm men chuyển gen biểu hiện ORF của chúng với sự có mặt của cơ chất PUFA. Kết quả của chúng tôi cho thấy khả năng kéo dài đối với PUFA được bổ sung ngoại sinh khác nhau giữa các elongase A. franciscana (Bảng 1). Đầu tiên, cả Elo2 và Elo5 đều có thể chuyển đổi tất cả PUFA đã được thử nghiệm (18:3n-3, 18:2n-6, 18:4n-3, 18:3n-6, 20:5n-3, 20:4n-6, 22:5n-3 và 22:4n-6) thành các sản phẩm kéo dài 2-cacbon tương ứng (Bảng 1). Hơn nữa, A. franciscana Elo1 và Elo6 có điểm chung là khả năng kéo dài tất cả các chất nền PUFA C18 đã được thử nghiệm cũng như 20:5n-3, với 22:5n-3 được Elo1 kéo dài thành 24:5n-3 chứ không phải Elo6 (Bảng 1). A. franciscana Elo3 và Elo4 chỉ công nhận hai PUFA C18 làm cơ chất, đó là 18:4n-3 và 18:3n-6 cho Elo3, và 18:2n-6 và 18:3n-6 cho Elo4, không phát hiện thấy hoạt động nào đối với Elo4. bất kỳ PUFA C20 và C22 nào (Bảng 2). Cuối cùng, A. franciscana Elo7 cho thấy khả năng kéo dài tất cả các chất nền PUFA C18 và C20 lên đến các sản phẩm PUFA C22, nhìn chung hoạt động mạnh hơn trên các PUFA n-3 so với PUFA n-6. Ngoài ra, một số elongase như Elo1 và Elo7 qua trung gian phản ứng kéo dài từng bước ngoài sản phẩm kéo dài đầu tiên (Bảng 1). Do đó, Elo1 đã kéo dài 18:3n-3 được cung cấp ngoại sinh thành 20:3n-3, sau đó được kéo dài thêm thành 22:3n-3, trong khi việc bổ sung 18:4n-3 và 18:3n-6 vào nấm men biểu hiện A .franciscana Elo7 đã dẫn đến sự tổng hợp các sản phẩm kéo dài C20 và C22 tương ứng.
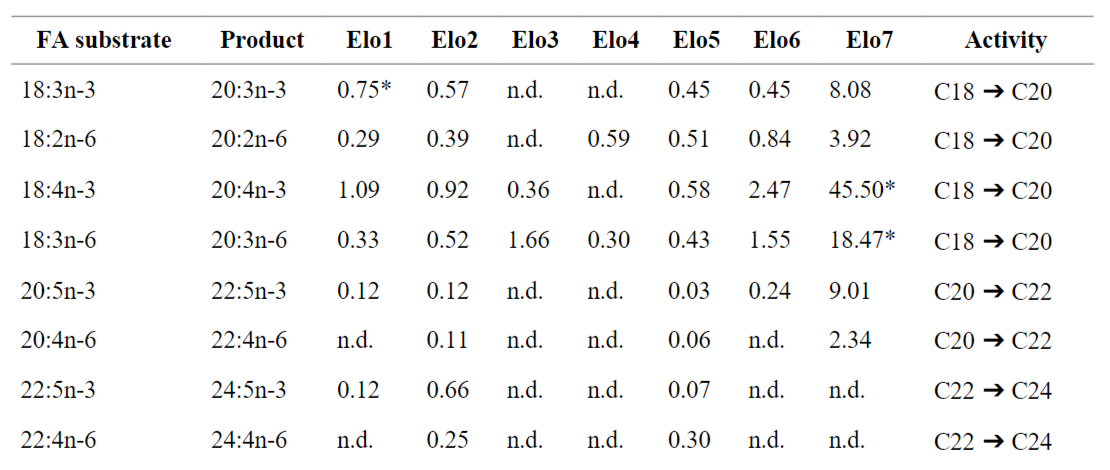
Bảng 1. Đặc tính chức năng của A. franciscana elongase. Chuyển đổi cơ chất axit béo không bão hòa đa (FA) được bổ sung ngoại sinh được tính theo công thức [diện tích sản phẩm/ (diện tích sản phẩm + diện tích cơ chất)] x 100.
Chức năng của A. franciscana desaturase
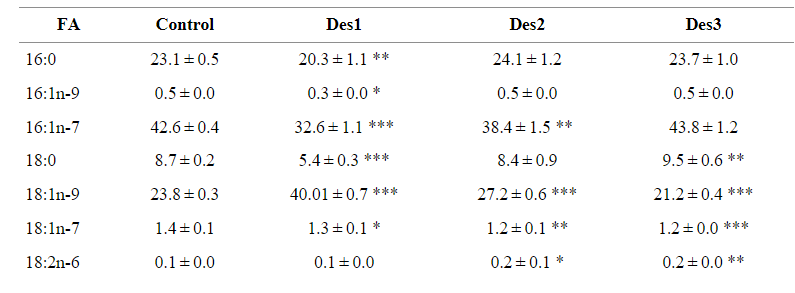
Hoạt động khử bão hòa của A. franciscana desaturase (Des1-3) đã được nghiên cứu bằng cách trồng nấm men chuyển gen biểu hiện ORF của chúng trong điều kiện không có chất nền FA được bổ sung ngoại sinh và khi có LA được cung cấp ngoại sinh. So sánh cấu hình FA của nấm men đối chứng và nấm men chuyển gen biểu hiện từng chất khử bão hòa của A. franciscana và được phát triển trong điều kiện không có bất kỳ chất nền FA bổ sung nào, cho thấy rằng nấm men biểu hiện A. franciscana Des1 và Des2 có hàm lượng ∆ cao hơn đáng kể 9 sản phẩm khử bão hòa 18:1n-9 (OA) (Bảng 2). Mặc dù chỉ có ý nghĩa đối với Des1, nhưng mức độ giảm của chất nền khử bão hòa ∆9 18:0 đã được phát hiện ở nấm men biểu hiện Des1 và Des2 so với nấm men đối chứng (Bảng 2). Cả Des2 và Des3 đều cho thấy mức tăng đáng kể 18:2n-6 (LA) so với nấm men đối chứng, cho thấy những gen này có hoạt tính ∆12 desaturase (Bảng 2). Cuối cùng, không có đỉnh bổ sung nào tương ứng với các sản phẩm khử bão hòa được phát hiện trong nấm men chuyển gen phát triển với sự có mặt của LA, cho thấy rằng bất kỳ desaturase nào thu được từ A. franciscana đều có hoạt tính ∆15 desaturase (dữ liệu không được hiển thị).
Tóm lại, các tác giả đã xác định được 8 elongase và 3 acyl desaturase béo từ A. franciscana. Trình tự suy ra của bảy trong số tám elongase được tìm thấy ở A. franciscana có đặc điểm của enzyme elongase PUFA. Các phân tích chức năng ở nấm men đã xác nhận A. franciscana có thể thực hiện tất cả các phản ứng kéo dài liên quan đến con đường sinh tổng hợp LC-PUFA, với Elovl8 (ở đây được gọi là “Elo7”) đóng một vai trò nổi bật rõ ràng. Tuy nhiên, kho các acyl desaturase béo có ở A. franciscana bị giới hạn ở ba gen được gọi là “các desaturase đầu tiên”, hai trong số đó (Des1 và Des2) có hoạt tính ∆9 desaturase dự kiến cho phép sinh tổng hợp 18:1n -9, trong khi Des3 thể hiện hoạt tính ∆12 cho phép sinh tổng hợp 18:2n-6. Cả chất khử bão hòa methyl-end và front-end đều không được tìm thấy ở A. franciscana. Nhìn chung, nghiên cứu hiện tại đã chứng minh rằng A. franciscana có khả năng sinh tổng hợp LC-PUFA hạn chế do thiếu các chất khử bão hòa chính trong bộ gen của nó và xác nhận rằng các quy trình làm giàu hiện tại đảm bảo cung cấp axit béo thiết yếu ngoại sinh cho ấu trùng biển là bắt buộc để ngăn chặn. triệu chứng thiếu hụt.

_1769663223.jpg)

_1769399561.jpg)
_1762138517.jpg)